 ОГЛАВЛЕHИЕ
>>>
ОГЛАВЛЕHИЕ
>>>
Как выше было сказано, я полностью исключил представление об ископаемых троглодитидах как охотниках. А ведь именно такое представление с необходимостью влечет приписывание им тех или иных свойств по аналогии с человеком. Мы имеем право утверждать, что троглодитиды даже и не могли бы убивать, ибо им это запрещал жизненный инстинкт – абсолютный, не допускающий исключений. Те популяции, которые нарушали бы эту биологическую норму поведения по отношению к животной среде, вымерли бы; иными словами, "не убивать" – это был наследственный безусловный рефлекс, врожденный видовой закон, безоговорочно закрепленный естественным отбором, а не навык, от которого особь могла бы и отвыкнуть. Теперь этот вывод нам важен и для того, чтобы нащупать едва ли не самое неясное и зыбкое звено во всей цепи анализа и реконструкции происхождения человеческой речи. Мы говорили, что невозможно вообразить себе семейство троглодитид в роли хищников, – у его представителей на всех уровнях не было почти ничего для нападения, а в природе все объекты нападения имеют те или иные средства самозащиты от хищников: рыбы уходят в воду, птицы взмывают в воздух, копытные убегают и т.д. Но теперь посмотрим оборотную сторону медали: сколь трудно представить себе троглодитида в роли нападающего, столь же трудно, оказывается, вообразить себе и защиту его самого от хищников. Что касается обезьян, они защищены древесным образом жизни (некоторые виды – наскальным). Несомненно, наши предки отчасти сохранили подобные элементы самообороны: будучи легче тех крупнейших хищников, которые стали бы на них систематически охотиться, они могли быстрее последних и в менее доступных местах передвигаться посредством лазания, благодаря наличию (сохранению от обезьян) рук, по ветвям или по крутым скалам. Но если развить эту мысль последовательно, выходит, что предки человека не могли спуститься на землю с деревьев (или скал), ибо они были бы съедены. Так и думал Энгельс: на низшей ступени дикости люди "жили, по крайней мере частью, на деревьях; только этим и можно объяснить их существование среди крупных хищных зверей".
Однако за прошедшие сто лет мы неоспоримо узнали, что они таки к тому времени спустились с деревьев. Значит, нужно как-то иначе объяснить их существование среди крупных хищников. Мысль археологов и антропологов искала разгадку лишь в одном направлении: в увеличении боевой силы наших предков в результате, с одной стороны, их вооруженности палками и камнями, с другой – соединенных действий группами. Это имеет некоторый филогенетический резон, так как обезьяньи стаи подчас успешно противостоят таким хищникам, как леопард. Но все это, даже если бы отвечало действительности, рисовало бы нам картину "оборонительного" приспособления к хищникам. А не было ли оно "наступательным", хотя и не в обычном смысле слова? Экологический анализ показывает нам колоссальную связанность палеоантропа со всем окружающим животным миром. И абсолютно иными путями, "палеонтологическим анализом языка", столько раз оспоренный и все же притягательный своим талантом и прозрениями лингвист Н. Я. Марр снова и снова возвращался к одному из своих казавшихся парадоксальными тезисов: наидревнейшие слои языка свидетельствуют о некоей тесной связи перволюдей с окружающим животным миром, какую нынешний человек не может себе и представить.
Не упускала ли до сих пор наука о происхождении человека из виду гигантские возможности активного воздействия высокоорганизованных предков человека на центральную нервную систему животных, на их высшую нервную деятельность? Если змеи "гипнотизируют" обезьян, то почему бы отказать высшим приматам в свою очередь в чем-либо подобном. У них степень подвижности нервных процессов выше, чем у других животных. Почему бы не применить это преимущество, не использовать слабые стороны нервной деятельности, поведения других видов. К сожалению, нигде не обобщены широко известные, но разрозненные сведения, что хищные не могут долго выдерживать взгляд человека. Не остаток ли это некоторой древней адаптации? Представим себе, что, еще не умея говорить между собой, троглодитиды могли адресовать каким-либо животным зримые или слышимые тормозные сигналы типа интердикции, которые в нашей сегодняшней речи преобразовались во что-нибудь вроде "киш", "фу", "брысь".
Только не упрощать! Конечно, палеоантроп не мог оказывать сигнального воздействия на все виды, на всех особей. Палеоантроп прежде всего укрывался от опасных видов тем, что использовал их природных антагонистов и конкурентов, стимулируя их враждебность и разобщение. Если в верхнем палеолите человек углубил антагонизм двух разновидностей волков – ныне дикого и предка собаки, а вместе с тем этого последнего со всеми другими хищниками, то от данного явления анализ может нисходить в глубь времен. Так, гиена отгоняла других от своей норы, и палеоантроп в какой-то мере, найдя те или иные успешно воздействующие на нее сигналы, был в некоторой безопасности "за ее спиной", по крайней мере пока находился в соседстве с ней. Впрочем, во множестве случаев это средство не годилось и приходилось больше рассчитывать на преимущество, которое давали собственные цепкие руки, – отсюда пристрастие и археоантропов и палеоантропов к обрывам, отвесным берегам и т.п. Но, возвращаясь к антагонизмам среди окружавших их животных, надо учесть не только антагонизмы между видами, но, может быть, еще больше – между индивидами. Широко известно, как сложны разделы охотничьих участков между особями того же хищного вида; " пристроившись" к одному, уже можно было не опасаться соседних.
В пользу самой возможности такого "сожительства" говорят разные данные полевых зоологов. Установление контактности исследователя с тем или иным хищным животным, даже на простом условнорефлекторном уровне, оказывается возможным. Так, в южнотаежном сибирском заповеднике "Столбы" сотрудница метеостанции настолько приручила одну из страшных хищниц – рысь, что та на зов своей "хозяйки" выбегает из чащи и сопровождает в экскурсиях по лесу не только ее, но и ее гостей. Опубликованы данные натуралистов о безопасном длительном проживании их и относительном контакте среди медведей, среди волков. Этот результат достигается только длительным и осторожным общением, однако высшая нервная деятельность человека неизменно берет верх. Кажется, особенно легко устанавливаются отношения полуприрученности с ирбисом (снежным барсом), который, впрочем, и вообще никогда не нападает на человека.
Попутно скажем, что в природе нет "диких" животных (ни хищных, ни травоядных) в обыденном смысле слова: в некоторых местах Африки непуганые животные разных видов, движимые ориентировочным инстинктом, приближаются к человеку, если только нет резких движений, которые вызовут иную реакцию – пассивно-оборонительную, т.е. бегство. Но в этом случае отрицательным раздражителем является не сам человек, а необычное резкое движение, хотя бы оно исходило и не от человека: если нет сильного ветра, заставляющего двигаться ветви и траву, животное убегает просто от какого-нибудь "необъяснимого" шевеления, способного таить опасность. Что касается хищников, нападение их на человека вообще мало характерно. Оно наблюдается там и со стороны тех видов, где имеет место постоянная охота, т.е. человек представляет опасность. Конечно, бывают особи-людоеды. Однако, как мало-мальски регулярная добыча, человек не только сейчас нигде не входит в нормальный пищевой рацион хищника, но и не входил на памяти истории, такие факты в биологии хищников всегда представляли отклонения от нормы. Надо только уметь обращаться с диким животным на воле, говорят натуралисты. Несмотря на зрелищную эффектность дрессировки хищников, она для биолога свидетельствует об их врожденной совместимости с человеком.
В отношениях человека с животными и в настоящее время дело отнюдь не сводится простой противоположности: дикие и домашние животные. Надо пользоваться и такими понятиями, как непуганость, полуприрученность и прирученность – по отношению к животным, которых искусственный отбор отнюдь не преобразовал в специальные одомашненные виды. Полуприрученными и прирученными являются, конечно, не виды, а индивиды, особи, в лучшем случае отдельные стада или стаи. Эта прирученность оказывается индивидуальным признаком, несколько противопоставляющим данного индивида (например, данную рысь в приведенном примере) прочим особям того же вида. Большее или меньшее приручение не индивидов, а стад (кабанов) или стай (волков) предполагают в мезолите. Одомашнением же называется возникновение новых видов, и оно датируется в основном временем неолита и позже. В настоящий момент в поле нашего зрения попадает как раз не видообразование, а диапазон явлений от непуганости до прирученности или, если угодно, симбиотичности, но не в экологическом смысле, а на уровне нейрофизиологических взаимодействий.
Впрочем, как проведешь жесткую границу между биологической взаимной пользой и нейрофизиологической адаптацией между индивидами разных видов? Лишь недавно выяснилось, что подчас бабуин в Африке выступал как бы в роли пастуха в стадах некоторых парнокопытных. Обезьяна издавала предупреждающие крики при возникновении опасности, подтаскивала заблудившихся детенышей при беспокойном крике самок в стаде, но зато, видимо, сама поедала их ослабевших или больных детенышей, можно предположить, что иногда и сосала молоко. Готтентоты издавна дрессируют отдельных бабуинов, используя их инстинкты и превращая в неплохих "пастухов" козьих стад.
По такому образцу мы можем представить кое-что и во взаимоотношениях ископаемых троглодитид со стадами травоядных. Если не усматривать предвзято в доисторическом прошлом обязательно войну нашего предка со всем животным миром, то откроется широчайшее поле для реконструкции его необычайно тесной и бескровной. связи с этим миром. Это, а не версия об охоте, важнейшая сторона процесса, который приведет его к порогу очеловечения. Даже в памяти первых европейских переселенцев в Южную Америку запечатлелось явление обитания вместе с семьями индейцев большого числа практически бесполезных разнообразнейших прирученных животных. Но тогда эти нравы и сцены, конечно, были не более чем реликтом.
В распоряжении биологической науки есть своеобразное средство для распознания, с представителями каких животных видов троглодитиды ранее всего и полнее всего находились в мирных взаимосвязях, во взаимном "приручении". Этим средством познания далекого прошлого ледниковой эпохи является удивительный факт весьма разной степени приручаемости и дрессируемости современных зверей и птиц разных видов. Конечно, правы те, кто утверждает, что разумное применение методики условнорефлекторной адаптации всегда дает некоторый положительный результат. Но бесспорно и то, что дрессировщикам в весьма разной степени удается нащупать у того или иного вида врожденную готовность к восприятию "антропического фактора" их поведения. По-видимому, практический опыт всякого рода зоопарков, зверинцев и цирков свидетельствует о худшей в общем приручаемости диких видов животных Нового Света (где не было ископаемых троглодитид) по сравнению с животными Старого Света. Вероятно, можно было бы построить целую иерархию всех животных по степени их на- / следственной приспособленности воспринимать тормозные или направляющие команды человека. Любопытно, что хищники Старого Света займут в этой иерархии далеко не последние места, а, скажем, гиены окажутся одним из самых "легких" объектов для цирковых дрессировщиков. Со временем с этой палеоэтологической точки зрения ученые рассмотрят и – ныне сенсационную – высокую дрессируемость дельфинов. Во всяком случае это очень перспективная научная дисциплина: восстановление ближайшей зоологической среды палеоантропов по коэффициенту дрессируемости и тем самым синантропичности всяческих видов животных, филогенетически восходящих к плейстоцену.
Если постараться представить себе, как это полуприручение и приручение могло протекать на практике, надо выдвинуть на первое место общение палеоантропа с детенышами животных, может быть, похищение их и вскармливание. Именно лабильность и адаптивность нервной деятельности в раннем онтогенезе животных представляли палеоантропу колоссальное многообразие и могущество средств их "воспитания". Для многих инстинктов в раннем онтогенезе еще не включены "пусковые механизмы", например подражание взрослым особям своего вида; поэтому эти инстинкты в раннем детстве могут быть полностью или частично угашены. С другой стороны, как мы уже знаем, имитативность особенно сильно действует в раннем возрасте. А став взрослыми, эти хорошо прирученные особи сохраняли, естественно, контактность, тормозимость, редуцированность некоторых инстинктов и могли служить буфером между себе подобными и палеоантропами.
Все сказанное – лишь подступ к проблеме: вскрыв явление и механизм интердикции (см. гл. 5, разд. VI), следует ли сразу переходить к зачаткам второй сигнальной системы в общении между людьми или нужен данный посредствующий блок – ив эволюционно-биологическом и в нейрофизиологическом смысле. Второе вероятно, но сегодня мы можем только предположительно говорить об этом и указать разрозненные симптомы из разных сфер знания, свидетельствующие в пользу возможного появления в будущем цельной концепции.
В современной человеческой речи в отличие от звуков, издаваемых животными, господствуют звуки, производимые струей выдыхаемого воздуха; животные, напротив, как правило, используют вдыхаемую струю. Но человек все же использует и этот прием "инспираторных" шумов. Я имею в виду не то исключение из общего правила, что в языках бушменов и готтентотов налицо немного инспираторных, в том числе щелкающих, звуков, подобных звукам подзывания животных, чмокания и т.п. Но дело в том, что и во всех человеческих языках такие инспираторные звуки используются в качестве междометий или в обращении к животным. Это дает основание считать такие звуки остатком древнейшей стадии. Следующий логический шаг, может быть, и ведет к представлению, что древнейшая "звуковая речь" адресовалась не от человека к человеку, а от человека (точнее – его предка) ко всевозможным иным животным. Ныне в обращении с животными мы употребляем не только эти оставшиеся от прошлого вдыхательные звуки, но и особые интонации, недопустимые по отношению к людям.
Наряду с такими частными звуками и интонациями замечено вообще явление реакции всевозможных животных на сам звук человеческого голоса – будь то речь или пение. Индейский писатель-натуралист канадец Вэша Куоннезин (Серая Сова) описал, как шаг за шагом он выработал некое специальное слово, произносившееся с одной и той же интонацией на определенной высоте звука. Автор пишет, что это слово приобрело "магическую силу" над зверями – оно их успокаивало, освобождало от тревоги. Если при его неожиданном появлении на виду у белок, мускусных крыс, бобров, лосей или при любом необычном звуке все они тревожно замирали, "словно каменные изваяния", то стоило произнести это уже привычное им всем слово (хоть звуки его были чужды зверям от природы), – все они, как один, оживали и продолжали прерванную жизнедеятельность.
Никак невозможно утверждать, что миф о пении Орфея, зачаровывавшем всех птиц и зверей, отразил какую-нибудь реальность. Но вот наблюдения из моего личного, куда более скромного опыта: когда я пел в лесу для собственного удовольствия, не раз птицы поднимали галдеж; когда однажды запел вблизи лошади, она ответила ржанием; когда пел в присутствии собаки (эрдельтерьер), она интенсивно подвывала. Интересно, что в особенности эти подражательные звуки вызывали самые высокие тона моего голоса.
Некоторым людям свойственно умение подражать голосам разных животных. Но лишь единожды был описан, а именно голландским врачом Тульпом в XVII в., "блеющий юноша", не обладавший ни в малой степени человеческой речью, но приспособившийся к блеянию, поскольку он вырос среди диких овец в горах Ирландии. Однако описание Тульпом особенностей морфологии его черепа делает более вероятным то, на первый взгляд совершенно невероятное, предположение, что это был вовсе не человек, а реликтовый палеоантроп. Если так, он мог "разговаривать" с животными, но еще не с людьми.
Все это – разрозненные клочки, которые невозможно пока соединить даже в сколько-нибудь цельную гипотезу. Пока назовем это лишь допущением. Подражая видовым голосам животных, в немалой части представлявшим собой неадекватные рефлексы, палеоантроп был вооружен сильным и небывалым оружием: он вызывал их имитативно-интердиктивную реакцию. В своем еще нечеловеческом горле он собрал голоса всех животных раньше, чем обрел свой специфический членораздельный голос. Итак, если наши реконструкции и допущения справедливы, палеоантроп занял совсем особое место в мире животных. Этот эврибионт, даже убиквист, т.е. обитатель неограниченно разнообразных биотопов, был абсолютно безопасен для всех зверей и птиц, ибо он никого не убивал. Но зато он как бы отразил в себе этот многоликий и многоголосый мир и смог в какой-то мере управлять поведением его представителей благодаря опоре на описанные выше механизмы высшей нервной деятельности.
Социальная психология как наука будет неполна, ибо не сможет вести нас в глубины истории и доистории, пока не включит в себя асоциальную психологию. Последняя должна состоять, очевидно, из двух разделов: а) криминальная психология (психология преступлений), б) патологическая психология (психопатология). Но криминальную психологию нам придется отложить: слишком много изменялось в истории в представлениях о "норме" и "преступлении". Революционер считался (и в капиталистических странах считается) преступником, его казнили, или ссылали, или держали в заключении. Джордано Бруно тоже был казнен как преступник. "Подрывные" мысли преследовались как преступление, хотя в другую эпоху они же вознесены как величайшая общественная ценность. Как показал французский исследователь Фуко, еще в XVII и XVIII вв. во Франции преступники и умалишенные содержались в одном заведении, так как сводились к некоему общему социальному знаменателю – ненормальному поведению, иными словами, нарушению принятых в данном обществе и в данное время норм социального поведения. Но мы обратим главное внимание на психопатологию как еще один источник, способный вести исследователя в глубины той дивергенции, с которой пошел род людской.
Психическое заболевание не установлено, пока нет ненормального поведения. Что же такое ненормальное поведение в самом широком обобщении? Это не те или иные действия, а невозможность их корректировать извне, т.е. привести в соответствие с требованиями среды или отдельных людей. Следовательно, ненормальность с точки зрения психологии – это невнушаемость. Такое V определение справедливо для любого общества, для любой эпохи. Что именно внушается, какие именно нормы поведения, речи и мышления – это исторически изменчиво. Но психическая болезнь состоит в нарушении элементарных механизмов, с помощью которых вообще люди подвергаются суггестии со стороны других людей (исключение составляют слабая олигофрения, т.е. дебильность, и микроцефалия, при которых внушаемость, напротив, гипертрофирована).
Болезнь ли это в точном смысле слова, т.е. имеются ли нервно-мозговые нарушения во всех случаях отклонения человека от нормального диапазона внушаемости? Еще сравнительно недавно все психические заболевания делили на две группы: органические и функциональные (чисто духовные). Только у первых поддавались наблюдению и определению те или иные нарушения в нервно-мозговом субстрате. Сюда относятся опухоли, повреждения кровоснабжения мозга, инфекции, интоксикации, травмы, врожденные аномалии морфологии мозга, нейрогистологические изменения. Функциональные же заболевания представлялись бестелесными. По поводу них возможны были философско-психологические спекуляции.
Но вот стена, разделявшая эти две группы, была пробита. Роль тарана пала на фармакологию: оказалось возможным "средствами химии лечить дух". Вернее, химия не лечит сам корень болезни, а компенсирует нечто, порождающее ненормальное поведение, подобно тому как издавна компенсируют близорукость и дальнозоркость с помощью очков или – с более недавнего времени – диабет с помощью инсулина. Но это "нечто", подправляемое химией, будет выглядеть как некоторое число разных явлений, пока мы не сумеем свести его к единому явлению – к проблеме внушения.
Однако, прежде чем прийти к такому обобщению, нужно сделать другой очень важный вывод из великих фармакологических побед над "нематериальными" психическими процессами. Раз химическими средствами можно воздействовать на самые различные формы ненормального поведения (как и обратно – делать поведение ненормальным), значит, непосредственная причина во всех этих случаях – нарушения в химизмах, в обменных процессах, осуществляемых тканями мозга. Но тем самым рушится принципиальное отличие от "органических заболеваний": химические процессы и их изменения и нарушения – это не менее органическое, т. е. "телесное", явление, чем поражения кровеносных сосудов головного мозга или структур нервных тканей.
Более того, сходство и даже общность многих симптомов при "органических" и "функциональных" психических болезнях заставляет считать, что и все структурно-морфологические изменения при "органических" болезнях ведут к ненормальному поведению больного через посредствующий механизм нарушения обмена веществ, т.е. через вызываемые ими сдвиги в химии тканей мозга (гистохимии). Таким образом, бурные успехи технологии фармакологических средств дали в руки психиатров возможность не только подлечивать психические болезни, но и доказать, что это бесспорно материальные нарушения, ибо то, что можно снимать введением в организм тех или иных химических веществ, несомненно, порождено отсутствием или недостатком в организме этих веществ (или иных, действующих аналогично).
Но речь идет не об одном веществе, а о весьма разных в том числе даже противоположных по своему действию. Так, например, одна группа медикаментов снижает неумеренную, буйную психическую активность, успокаивает, а в больших длительных дозах угнетает, приводит к депрессии, сонливости и летаргии. Другая же группа преодолевает психическую пассивность, унылость, меланхолию, депрессию, но при больших длительных дозах превращает недоактивность в сверхактивность, подавленность – в буйство. Одни химические агенты корректируют чрезмерную активность, другие – недостаточную. Есть и более специальные медикаменты.
Что же объединяет все эти разные явления? Только то, что и повышенная и пониженная активность делают человека в той или иной мере неконтактным и асоциабильным. Это значит, что окружающие не могут в должной мере влиять на его поведение. Вот почему медикаментозная терапия (химиотерапия) – всегда сочетается врачами с психотерапией, в том числе с мягким, заботливым обращением, восстанавливающим разрушенные или недостающие мостки контактности и открывающим дорогу внушению в широком смысле (в том числе, если надо, и гипнотическому). Неконтактность – это и есть броня, закрывающая психотика от внушения окружающих. Неконтактность тождественна невнушаемости. И в самом деле, эту функцию в равной мере выполняют обе противоположные аномалии: если психотик сверхактивен, он заблокирован от воздействия слов и поступков других собственными маниями (стойкими самовнушениями), бурной двигательной активностью или, наоборот, кататонией, которые невозможно перебить никаким внушаемым, т.е. требуемым, рекомендуемым, испрашиваемым, действием; если психотик слабоактивен, он заблокирован от воздействия слов и поступков других своей нереактивностью, депрессивностью, дремотой. Оба противоположных фильтра схожи, так как в равной мере не пропускают тех же самых воздействий внушения: один -неукротимость, другой – недоступность. Иными словами, оба характеризуются неполной проницаемостью или даже непроницаемостью для специально антропических раздражителей. Один – маниакальное упорство, другой -капризность. Следовательно, нормальный человек, т.е. поддающийся и подвергаемый внушению, вернее, идущий навстречу внушению, находится в узком диапазоне между этими двумя крайностями (оставляя здесь в стороне явление гипервнушаемости). Это как бы щель в спектре невнушаемых состояний, точка уравновешенности двух возможных противоположных по своему знаку состояний невнушаемости. Недаром при современном медикаментозном лечении в любом случае прописываются оба противоположно действующих средства в разных пропорциях, чтобы предотвратить прямой перевал из одного невнушаемого состояния в противоположное, не удержавшись в критическом переломном интервале. Если не применять комбинированных медикаментов, именно это и получается.
Подойдем к этим явлениям с антропогенетической точки зрения. Все психические заболевания теперь придется поделить на две совсем новые группы: генетически обусловленные (маниакальные и депрессивные психозы, олигофрении, шизофрении и т.д.) и экзогенные (травматические, наркотические, токсические, инфекционные, опухолевые). Нас интересует только первая группа, вторая же лишь постольку, поскольку она способна воспроизводить частично или вполне симптомокомплексы первой. Несмотря на бурное развитие генетики, еще почти никто не подошел с позиции антропогенеза к материалу психиатрии.
Правда, до появления и генетики, и психофармакологии уже зародилось научное направление такого рода, но крайне узкое. Это – эволюционный подход к микроцефалии, связанный с именами Фохта (1868 г.) и Домбы (1935 г.). Мысль была правильная: некоторые врожденные психические аномалии представляют собой атавизмы, т.е. возрождение в редких особях того, что было всеобщим в филогенетически предковой форме. Такой атавизм Фохт и Домба усмотрели в симптомокомплексе микроцефалии. Это было очень демонстративно и истинно. Но, во-первых, они были вынуждены ограничиться только той аномалией, при которой налицо прежде всего выраженные физические уклонения от нормы: малоголовостью, морфологически "предковыми" признаками черепа (и мозга), а уж глубоким слабоумием только как сопровождающим синдромом. Во-вторых, они не, могли опираться на генетику, т.е. научно объяснить и неизбежность возрождения в потомстве предковых черт, и в то же время неизбежность расщепления предковых черт, т.е. невозможность повторения полного портрета предковой формы среди особей последующей биологической формы. В-третьих, ввиду недостаточного развития антропологии – не только при Фохте, когда ее почти вовсе не было, но и при Домбе, когда она немало продвинулась, – они не имели достаточного понятия о тех предковых родах, из которых произошел Homo sapiens, т.е. об археоантропах и палеоантропах.
Но вот сегодня мы можем значительно обобщить открытие Фохта – Домбы: все истинные или генетически обусловленные психические болезни можно считать воспроизведением разрозненных черт, характеризовавших психонервную деятельность на уровне палеоантропов или, крайне редко, более отдаленных предков. Ведь генетически обусловленными могут быть не только морфологические и морфофункциональные, но и обменные, гистохимические, химико-функциональные отклонения от нормы у неоантропа в сторону палеоантропа. Последние, т.е. обменные отклонения в тканях мозга, мы обнаруживаем только пр. ненормальному поведению. И в самом деле, среди характерных аномалий поведения душевнобольных сколько замечаем мы признаков, которые ныне исследователи реликтовых гоминоидов (палеоантропов) описывают как свойства последних. Например, ночное блуждание (лунатизм), летаргия и длительный неглубокий сон или дремотное состояние, гебефрения – беспричинный смех и ряд других.
Но нам сейчас важен один, причем негативный признак всех психических патологий: они воспроизводят эволюционную стадию невнушаемости, т.е. не контрсуггестивность, а досуггестивность. Впрочем, здесь есть элемент оборонительной функции, как бы забронированность от суггестивной (или, может быть, лишь интердиктивной) работы возникающей второй сигнальной системы. Видимо, это как раз и восходит к нейропсихическим чертам палеоантропов эпохи дивергенции.
Дело не в содержании тех или иных маний и бредов: воображает ли себя больной Христом, Наполеоном или Гитлером, воображение тут в любом случае подкрепляет или оформляет его психическое состояние закрытости для внушения с чьей бы то ни было стороны ("выше всех"). Существенна и стабильна только эта функция, а не исторически случайная личина: уподобление себя великому человеку облегчает невнушаемость, а не является ее конечной причиной. Психопатология сверхактивности и слабой активности имеет тот же общий признак: "защищенность" от внушения; я ставлю это слово в кавычках, так как внушение вовсе не обязательно отождествлять с причинением ущерба, оно вполне может играть и обратную роль. Вот почему это свойство палеоантропов правильно называть досуггестивным – они были просто еще вне социальных контактов, еще не обладали собственно второй сигнальной системой даже в зачаточной форме.
Из сказанного есть, как выше отмечено, лишь совсем немного исключений: микроцефалы и дебилы обладают повышенной внушаемостью; криминалистике известно, что преступники широко используют дебилов как свое орудие, злоупотребляя их внушаемостью. Дебильность – самая слабая степень олигофрении. Но ведь суггестивная функция еще не есть зрелая вторая сигнальная система, она принадлежит кануну или началу второй сигнальной системы. Тем самым эта сверхвнушаемость при некоторых прирожденных психопатиях все же свидетельствует о том, что на каком-то этапе или в какой-то части популяции было нормой в мире поздних палеоантропов или, может быть, ранних неоантропов. Мы можем даже предположить, что в их отношениях боролись между собою эти две тенденции: асуггестивность и гиперсуггестивность.
Итак, психически больные люди – это неизбежное, по законам генетики, воспроизведение в определенном маленьком проценте человеческих особей отдельных черт предкового вида – палеоантропов. Речь идет ни в коем случае не о широком комплексе, тем более не о полноте черт этой предковой формы, а лишь о некоторых признаках, самое большее – о группе необходимо коррелированных признаков. Точно так же у других человеческих особей воспроизводится, скажем, обволошенность тела без всяких других неандерталоидных симптомов, у иных – некоторые другие черты морфологии. Совсем попутно отметим, что вследствие существенно иного генетического механизма отдельные неандерталоидные признаки проявляются в старости, причем у женщин статистически несколько чаще.
Те люди, у которых в сильной форме воспроизводятся некоторые нервнопсихические черты предковой формы, попадают в категорию психически больных, т.е. в ведение психиатрии. Как мы уже обобщили, это в основной массе так или иначе невнушаемые (неконтактные) личности. С точки зрения норм нашей человеческой жизни это очень большое несчастье. Но нас интересует их симптомокомплекс лишь как памятник жизни существ, еще не перешедших в людей: психотики в условиях клиники или дома, загородившись от внушения, тем вынуждают обслуживать себя. Эти индивиды как бы вырываются из сети внушений, заставляющих людей действовать не по стимулам первой сигнальной системы и животного самосохранения. Они не могут умереть, ибо окружены заботой других. В положении психотика, таким образом, есть нечто генетически напоминающее паразитизм при вполне здоровом теле.
В мире животных нет психопатологии. Неврозы во всем предшествовавшем изложении сознательно элиминировались. Но и неврозы у животных могут быть только экспериментальными, т.е. в искусственно созданных человеком условиях. В природной обстановке животное-невротик было бы обречено на быструю гибель.
Целая цепь ученых от Уоллеса до Валлона доказывала и доказала, что человеческое мышление не является линейно нарастающим от животных предков полезным свойством; напротив, оно и в антропогенезе, и в онтогенезе у ребенка сначала вредно для каждого индивидуального организма, делает его беспомощнее по сравнению с животным; лишь дальнейшее его преобразование понемногу возвращает ему прямую индивидуальную полезность. Но как же, если исключить всякую мистику, объяснить это "неполезное" свойство? Ведь естественный отбор не сохраняет вредных признаков, а нейтральным признаком данное свойство не назовешь. Возможно лишь одно объяснение: значит, оно сначала было полезно не данному организму, а другому, не данному виду (подвиду, разновидности), а другому. Следовательно, надо изучить, во-первых, кому и почему это свойство у других было полезно, во-вторых, как они, заинтересованные, это свойство других закрепляли, удерживали, навязывали.
Мы не можем с помощью сказанного в этом разделе восстановить точную схему дивергенции троглодитид и гоминид, начавшуюся еще в мире поздних палеоантропов н завершившуюся лишь где-то при переходе от ископаемых неоантропов к современным. Мы можем лишь совершенно предположительно допустить, что поздние мустьерцы, в высочайшей мере освоив сигнальную интердикцию в отношении зверей и птиц, наконец, возымели тенденцию все более распространять ее и на себе подобных. Эта тенденция в пределе вела бы к полному превращению одних в кормильцев, других в кормимых. Но с другой стороны, она активизировала и нейрофизиологический механизм противодействия: асуггестивность, неконтактность.
Моя задача состоит не в предвосхищении этих будущих палеопсихологических исследований, а лишь в постановке наряду с предыдущими и этой части проблемы дивергенции троглодитид и гоминид. Это слишком ответственная задача, чтобы осмелиться на нечто большее, чем первый шаг.
Антропологи уже вполне удовлетворительно выяснили анатомо-морфологическую эволюцию человека. С их точки зрения, достаточно установить с помощью сопоставления скелетов, что неоантроп развился из палеоантропа, последний – из археоантропа (питекантропа) и т.д. Для них даже удобно, если это эволюционное древо рисуется не ветвистым, а прямым, как корабельная мачта: ведь им надо знать только, кто из кого произошел; предка можно посчитать исчезнувшим с того момента, как появился потомок.
Однако, поскольку эта работа в основных чертах выполнена, главной проблемой антропогенеза уже становится не морфологическое отличие неоантропа от предковой формы, а его жизненные отношения с ней. Человек не мог не находиться в тех или иных отношениях с видом, от которого он постепенно стал отличаться и отдаляться. Это были отношения экологические или биогеографические, отношения конкуренции, или симбиоза, или паразитизма, или какого-либо еще типа. Ведь различия углубляются лишь в процессе дивергенции разновидностей, поначалу же они незначительны. Наука об антропогенезе, думается, должна, наконец, стать наукой о конкретных биологических отношениях людей и той предшествовавшей формы, от которой они ответвились. Научной несообразностью является взгляд, будто все особи предкового вида превратились в людей. Еще бессмысленнее думать, что они перестали рождаться на свет с тех пор, как некоторые путем мутации стали людьми. Не лучше и идея, что немногие, ставшие людьми, в короткий срок лишили кормовой базы всех отставших и те быстро перемерли: на земле до сих пор остается довольно пищевых ресурсов для множества видов животных.
Все эти несуразицы только подчеркивают неоправданность упорного избегания темы о реальных взаимоотношениях двух разновидностей, вероятно, лишь в ходе этих взаимоотношений ставших подвидами, а затем и разными видами, продолжая и на этом таксономическом уровне находиться в биологических отношениях друг с другом. Таким образом, к науке об антропогенезе предъявляется пожелание перенести, наконец, главное внимание с вопроса об отличии людей от их ближних биологических предков на вопрос о реальных отношениях людей с этой предковой формой.
Мы близимся в науках о человеке к такому сдвигу, который можно сравнить с революцией в физике, развернувшейся в первой половине XX в. Роль, аналогичную "атомному ядру", здесь сыграет начало человеческой истории. Но сегодня это еще только штурмуемая загадка. Если принять, что все сказанное выше об экологии троглодитид более или менее соответствует истине, то начало человеческой истории круто переносится во времени в сравнении с принятой сейчас датировкой. Еще недавно длительность истории определяли в полмиллиона – миллион лет, и уже эта цифра в известной мере оправдывала тезис А. Тойнби, что сравнительно с нею история всех вычлененных им "цивилизаций" (числом около двадцати) настолько кратковременна, что последовательностью их можно пренебречь и рассматривать их почти как одновременные друг другу, т.е. не имеющие соизмеримости с гигантской величиной бытия людей на земле. Однако с тех пор раскопки Лики, Арамбура, Коппенса и других в Африке увеличили ее еще значительно больше, так что сегодня людям приписывают возраст около двух с половиной миллионов лет и, судя по всему, завтра могут последовать новые открытия еще более древних костных останков австралопитеков в сопровождении примитивных оббитых камней. Но вот что касается неоантропа (Homo sapiens), он появляется всего 35–40 тыс. лет тому назад. Его исторический марш, обгоняющий темпы изменения окружающей природы, т.е. обретающий относительное самодвижение и ускорение (при неизменности телесной организации), начинается и того много позже. Следовательно, при изложенных представлениях исторический процесс радикально укорачивается. Если отсчитывать начало такового самодвижения с неолита, эти недолгие тысяч восемь лет человеческой истории по сравнению с масштабами биологической эволюции можно приравнять к цепной реакции взрыва.
История людей – взрыв. В ходе ее сменилось всего несколько сот поколений.
Толчком к взрыву, очевидно, послужила бурная дивергенция двух видов – палеоантропов (троглодитов) и неоантропов, стремительно отодвигавшихся друг от друга на таксономическую дистанцию подвидов, видов, родов, семейств, наконец, на дистанцию двух различных форм движения материи – биологической и социальной.
Именно природа этой дивергенции и есть "атомное ядро", тайну которого надлежит открыть. Для начала анализа ясно лишь, что, будучи процессом биологическим, она в то же время имела нечто отличающее ее от всякой другой дивергенции в живой природе. К тому немногому, что мы достоверно об этом знаем, принадлежит необычная быстрота данного ароморфоза – отпочкования нового, прогрессивного вида. Отсюда можно сделать вывод, что между обоими дивергирующими видами должны были существовать и крайне напряженные экологические отношения. Этого не было бы, если бы дивергенция с самого начала сопровождалась размежеванием ареалов. Вероятнее, напротив, что в пределах общего ареала происходило крутое размежевание экологических ниш.
Но главный вывод, который мы должны извлечь из стремительности дивергенции (а ее отрицают только немногие антропологи вроде А. Валлуа, держащиеся слабо обоснованной концепции происхождения Homo sapiens не из того или иного вида палеоантропов, а из "пресапиенсов", восходящих к среднему или даже раннему плейстоцену), состоит в том, что перед нами продукт действия какого-то особого механизма отбора.
Прежде всего хотелось бы реконструировать не только само раздвоение или, вернее, отпочкование, но и его более мелкие промежуточные уровни. Мыслимо ли это?
Но следует ли вообще думать, что палеонтология всегда ищет и находит все промежуточные ступеньки между одной биологической формой и филогенетически последующей, уже существенно отличающейся? Состоит ли сама идея палеонтологии в том, что в принципе должны где-то существовать останки всех мыслимых степеней сочетания прежнего и нового? Нет, конечно, в этом филогенетическом переходном мосту всегда много неустойчивых, хрупких образований, не надстраивающихся в чисто количественном ряду друг над другом, а представляющих очень бедные по числу, очень вариативные и очень ломкие образования. Пока, наконец, одно из них не станет основанием для жизнеспособной, многочисленной ветви.
Палеонтологи иногда называют это практически неведомое им, исчезнувшее соединение эволюционных форм "черешком". Этот черешок, на котором держится новый вид, всегда тонок, почти никогда не доступен прямому изучению палеонтологии. Иначе говоря, в диапазоне между родительскими и нашедшими свою почву стойкими, дающими богатые соцветия таксономическими единицами находится обвал возникавших и гибнувших нежизнеустойчивых форм. В десятки раз труднее изучить этот "черешок" ответвления человека – Homo sapiens, оторвавшегося относительно быстро на огромную, как мы уже знаем, биологическую дистанцию: на расстояние нового семейства. Уж очень специфично то, что возникло: вид, отличающийся инверсией процессов высшей нервной деятельности, – "животное наоборот". Посмотрим, что же мы все-таки имеем в руках из костного материала, годного для непосредственной датировки и биологической фиксации дивергенции.
В результате блестящих исследований ископаемых эндокранов, осуществленных В. И. Кочетковой, мы узнали нечто более важное, чем существование тут и там в четвертичных отложениях "переходных" черепов, расположенных по сумме признаков на том или ином отрезке пути между "неандертальцем" и "кроманьонцем". Открытие Кочетковой состоит в глубоком изменении прежнего представления о самих кроманьонцах, т.е. об ископаемых неоантропах начальной поры верхнего палеолита, которые оказались не тождественными позднейшим неоантропам. Трудно переоценить огромность этого, казалось бы, тончайшего сдвига: кроманьонцы – не то, что привычно и долго о них воображали. А именно было общепринято, что кроманьонцы – это другое наименование для нас самих. Посади с нами за обеденный стол неандертальца – все согласны, что его общество было бы невыносимо; но посади кроманьонца (хорошо одетого, побритого, обученного нашему языку и манерам) – его якобы никто бы и не отличил. Соответственно подчас говорят: "Мы, кроманьонцы". Исследование эндокранов обнаружило тут ошибку. Трудно сказать, оценила ли в полной мере сама В. И. Кочеткова всю капитальность своего вывода, что черепа группы ископаемых неоантропов (Homo sapiens fossilis) серьезно отличаются – по крайней мере в некотором проценте экземпляров и тем самым в среднем – от величин типичных и устойчивых для ныне живущих неоантропов, т.е. людей современного типа. Мало того, выяснилось, что это отклонение характеризует людей первой половины верхнего палеолита (столь же неточно в общежитии именуемой "ориньяком"). Такие верхнепалеолитические индивиды, как Кро-Маньон III, Маркина Гора, оказались по эндокрану, т.е. по макроморфологии головного мозга, вообще ближе к палеоантропам, чем к неоантропам.
В своих цифровых таблицах различных параметров строения мозга В. И. Кочеткова убедительно выделила ископаемых неоантропов в особую группу, оказавшуюся глубоко специфичным перевалом в антропогенезе. Некоторые показатели, нарастающие во всей цепи от шимпанзе к австралопитекам и далее, достигают своей кульминации именно в группе ископаемых неоантропов, после чего кривая падает. Другие показатели, наоборот, достигают кульминации накануне появления этой группы, т.е. у палеоантропов, а с ископаемых неоантропов уже начинается нисходящая линия, характерная для неоантропов вообще по сравнению с ростом соответствующей кривой у троглодитид вообще. Однако следует помнить, что вся группа ископаемых неоантропов пока представлена сравнительно немногочисленными находками. Тем выразительнее выступает ее полиморфность (см. составленную мною сводную таблицу по опубликованным данным Кочетковой).
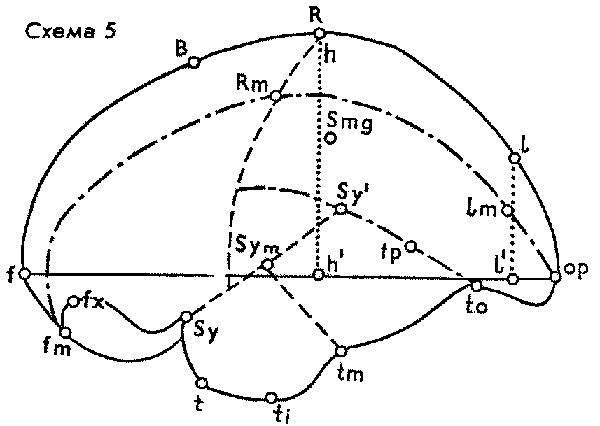 Таблица 1. Сводная таблица по
материалам В.И.Кочетковой (все цифры в мм, кроме двух
последних строк; величины средние, в скобках – минимум и
максимум; условные обозначения см. по схеме мозга)
Таблица 1. Сводная таблица по
материалам В.И.Кочетковой (все цифры в мм, кроме двух
последних строк; величины средние, в скобках – минимум и
максимум; условные обозначения см. по схеме мозга)
|
Понгиды |
Троглодитиды |
Гоминиды | ||||||
|
Шимпанзе |
Австра |
Археоантропы |
Палеоантропы |
Ископаемые неоантропы |
Современные люди | |||
|
Тотальный продольный размер |
106 (104-109) |
125 |
153,7 (145-159) |
174,7 (163-182) |
186.3 (176-196) |
167,1 (150-183) | ||
|
Лобная доля |
Верхний продольный диаметр fm – R |
81,3 (78-86) |
92,0 |
110,0 (98-118) |
128,7 (123-139) |
135,3 (98-147) |
129,4 (116-143) | |
|
Нижний продольный диаметр fm – Sy |
33,6 (31-35) |
55,5 (51-60) |
61,4 (58-65) |
55,0 (46-60) |
49,0 (43-57) | |||
|
Дуга нижнего лобного края fm – Sy |
56.3 (52-63) |
70,9 (61-79) |
83,8 (75-91) |
74,0 (71-76) |
70,2 (58-87) | |||
|
Дуга латериального бугра fx–Sy |
28,0 (22-34) |
41.5 (35-50) |
52.9 (46-58) |
44,4 (39-50) |
43,8 (41-47) | |||
|
Максимальный широтный размер р–р |
69,3 (66-75) |
89,0 (81-93) |
108,1 (103-115) |
108,0 (102-113) |
104,4 (89-126) | |||
|
Теменная доля |
Верхняя |
Продольный диаметр R-1 Высотный диаметр калотты h–h' |
31.3 (26-36) 48,0 (44-51) |
35,0 54,7 |
48,0 (48-51) 66,0 (61-69) |
57,5 (52-64) 77,7 (72-82) |
67,9 (60-76) 76,6 (68-90) |
65,8 (51-77) 85,9 (72-97) |
|
Нижняя |
Продольный диаметр Rт–1т Поперечный диаметр smg–smg |
40,3 (33-57)
71 (61-80) |
49,0 |
67,7 (65-70)
103,3 (94-112) |
82,3 (74-91)
118.9 (104-132) |
95,1 (87-101)
116.5 (101-140) |
88,1 (79-101)
124,1 (108-152) | |
|
Височная доля |
Продольный диаметр t-tp Поперечный диаметр eu–еu Высотный диаметр Sy–ti Высотный диаметр Sy'-to Высотный диаметр tm–Sym |
62,6 (61-65) 88,0 (86-89) 23,3 (21-25) 32,6 (31-34) 2 3,3 (21-23) |
66,0 |
73,5 (73-74) 123,3 (118-129) 34,0 (28-40) 33,5 (33-34) 32,0 (29-35) |
86,6 (78-94) 136,3 (116-146) 39,2 (34-46) 42.2 (36-46) 36,2 (32-41) |
92,5 (90-95) 136,3 (126-146) 47,0 (43-51) 51,0 (46-5S) 37,4 (33-42) |
90,4 (76-100) 132.2 (117-158) 42,7 (33-51) 46.2 (38-54) 42,0 (36-51) | |
|
Затылочная доля |
Продольный диаметр 1m–om Высотный диаметр 1-1' |
38,6 (35-41) 25,0 (22-27) |
38,0 37,0 |
42,3 (38-47) 44.0 (38-50) |
44,9 (39-50) 50,9 (44-59) |
42,9 (35-55) 50,6 (37-56) |
38,0 (29-52) 47,9 (39-54) | |
|
Степень нависания затылка над мозжечком (в относительных величинах) |
3,5 (2,9-3,8) |
6.8 (6.3-7.3) |
10,9 (9.3-13,5) |
8,9 (5,6-13,5) |
5,4 (2,9-9,5) | |||
|
То же (в баллах) |
0,7 (0.6-0,8) |
1.4 (1,3-1,5) |
2,1 (1,6-2.7) |
1,8 (1,2-2,7) |
1,1 (0,6-1.9) | |||
Из этой таблицы вполне правомерно вывести заключение, что ископаемые неоантропы - это и есть "черешок" нового семейства. Вернее, это пестрый конгломерат не очень жизнеспособных видов и разновидностей, составлявших переходный мост между палеоантропами и неоантропами современного типа, тем самым между двумя семействами. На дне пропасти между ними найдены лишь немногие обломки этого филогенетического моста. В переводе на хронологию его длина - всего лишь 15-25 тыс. лет. Но на этом-то отрезке и укладывается почти все таинство дивергенции, породившей людей. Впрочем, начало его надлежит продвинуть несколько дальше в прошлое: первый пролет моста кое в чем начинает вырисовываться в гуще поздних палеоантропов. Часть этих животных, как отмечено выше, уже обладала странностями вплоть до размазывания пятен красной охры - пережженной глины или окислов железа (эта странность - не "искусство", вспомним, что самец птицы австралийский атласный беседочник раскрашивает внутренность своей беседки, пользуясь кусочком предварительно измочаленной коры, - это чисто этологическое приспособление для отличения самками партнеров своего вида, исключающее межвидовое скрещивание). Мы уже знаем, что родовым, всеобщим отличительным свойством семейства гоминид, постепенно отходившего от троглодитид поэтому мосту, или черешку, является вторая сигнальная система. Следовательно, для всех представителей этой "переходной" группы может быть характерной выраженность разных компонентов, из которых вторая сигнальная система сложится в дальнейшем, однако пока в разрозненном виде еще не дающих устойчивой и жизнеспособной функции и структуры. Может быть, иные из этих компонентов выгодны одним особям и одновременно гибельны для других или выгодны особи в данный момент и гибельны в другой.
Таким амбивалентным компонентом могла быть описанная нами выше нейрофизиологическая (если угодно, палеоневрологическая) функция интердикции или надстраивающаяся над нею функция суггестии. Ведь пока эта последняя не породит из себя функцию контрсуггестии, хотя бы в ее зародышевых проявлениях, не может еще быть сколько-нибудь стойкой биологической или социальной системы. Интердикция, суггестия – это мощные факторы межиндивидуальных воздействий, но и порождающие, и снова разрушающие сами себя. Вот все эти преобразования от уровня интердикции до порога контрсуггестии, все эти чрезвычайно сложные и далеко еще не выясненные палеоневрологические трансформации и приходятся в основном на филогенетический интервал, о котором идет речь. Эти преобразования, вероятно, составят предмет долгих будущих исследований. Хорошо уже то, что мы можем указать и хронологические рубежи, в которые они вписываются, и их главное направление. Кончилось время, когда внимание палеоантропологов было роздано более или менее равномерно костным останкам наших двуногих предков, находимым на геологических глубинах до двух с лишним (или до четырех?) миллионов лет давности. Даже тем научно значимее представлялись эти останки, чем они залегали глубже, чем были древнее. Конечно, мы будем заниматься ими и впредь, так же как и ископаемыми высшими обезьянами, но проблема антропогенеза в точном и узком смысле теперь сфокусировалась на сравнительно недолгом интервале времени, но крайне насыщенном. Отныне надолго "загадка человека" будет всасываться в эту небольшую воронку – в неисчерпаемо сложную тему дивергенции палеоантропов и неоантропов.
Каковы же наши опорные знания сегодня о фактах, имевших место в этом интервале? К фактам, касающимся трансформации черепа и мозга и свидетельствующим о генезисе второй сигнальной системы, мы обратимся в последней главе. Сейчас существенна общая констатация: на протяжении этого отрезка макроморфология мозга еще менялась, позже – не менялась. А пока отметим лишь некоторые точно установленные факты более внешнего порядка.
В этом интервале в числе остатков жизнедеятельности наших ископаемых предков появляются сначала краски, в конце – изображения. Но как мустьерское использование охры для пятен на камнях, для отпечатков пятерни, так же и ориньякско-солютрейские насечки и полоски, графические и скульптурные изображения животных и людей, – все это не имеет ни малейшего отношения к категориям эстетики и отвечает столь ранним ступеням подготовки специфической человеческой психики, что эти явления должны быть поставлены в порядке эволюции у самых истоков возникновения речи. И все-таки тут налицо нечто высоко специфичное для становления человека: если и мыслимо животное, которое применяет элементарную окраску, то ни одно животное не создает изображения чего-либо.
Кроме того, есть и еще один совершенно специфический факт, который мы можем локализовать в данном хронологическом интервале: расселение ранних неоантропов по обширной ойкумене, чуть ли не по всей пригодной к обитанию территории нашей планеты, включая Америку, Австралию, Океанию. Эта дисперсия человечества по материкам и архипелагам земного шара, если сравнить ее с темпами расселения любого другого биологического вида, по своей стремительности может быть уподоблена взрыву. За эти полтора-два десятка тысячелетий кроманьонцы преодолели такие экологические перепады, такие водные и прочие препятствия, каких ни один вид животных вообще никогда не мог преодолеть.
Нельзя свести это рассеяние людей по планете к тому, что им не доставало кормовой базы на прежних местах: ведь другие виды животных остались и питаются на своих древних ареалах нередко и до наших дней – корма хватает. Нельзя сказать, что люди в верхнем плейстоцене расселялись из худших географических условий в лучшие, – факты показывают, что имело место и противоположное. Им не стало "тесно" в хозяйственном смысле, ибо их общая численность тогда была невелика.
Но им стало, несомненно, тесно в смысле трудности сосуществования с себе подобными. Старались ли они отселиться в особенности от палеоантропов, которые биологически утилизировали их в свою пользу, опираясь на мощный и неодолимый нейрофизиологический аппарат интердикции? Или они бежали от соседства с теми популяциями неоантропов, которые сами не боролись с указанным фактором, но уже развили в себе более высокий нейрофизиологический аппарат суггестии, перекладывавший тяготы на часть своей или окрестной популяции? Вероятно, и палеоантропы, и эти суггесторы пытались понемногу географически перемещаться вслед за такими беглецами-переселенцами. Но остается очень убедительным вывод современного расоведения: американские неоантропы-монголоиды (индейцы) по своему антропологическому типу древнее современных азиатских, т.е. откочевали из Азии в Америку до сколько-нибудь плотного заселения Азии, а из американских южноамериканские древнее североамериканских; австралийские аборигены представляют особенно древний тип неоантропов, т.е. переселились сюда в весьма раннюю пору формирования неоантропов. Из этих фактов умозаключение однозначно: на самые далекие края пригодного к обитанию мира неоантропы отселились особенно рано в эпоху дивергенции с палеоантропами. А судя по тому, что расселение ранних неоантропов происходило в особенности по водным путям – не только по великим рекам, но и по океанским течениям, на бревнах, – люди искали отрыва сразу на большие дистанции, передвигались они при этом, конечно, поодиночке или очень небольшими группами.
Но вот процесс разбрасывания то в том, то в ином направлении достигает такого предела, когда по природным причинам простое взаимное отталкивание оказывается уже далее невозможным. Достигнуты ландшафтные экстремальные условия, или океан останавливает перемещение дальше вперед. Но торможение может быть и иного рода: настигают новые волны человеческой миграции, отрываться все труднее. И вот рано или поздно в разных местах не в одно и то же время, но в общем повсюду приходит пора нового качества: взаимного наслаивания мигрирующих популяций неоантропов, откуда проистекают попытки обратного, встречного переселения. Теперь люди все чаще перемещаются не в вовсе необжитую среду, а в среду, где уже есть другие люди, пусть и редкие, где земли, растительности и живности хватает, но где необходимо как-то пребывать среди соседей. Иссякает отлив, начинается прилив. Люди возвращаются к людям. Или – что равнозначно – они уже не отселяются, они остаются среди людей.
Вот этот второй, обратный вал перемещений неоантропов и есть уже не просто история их взаимного избегания или избегания ими палеоантропов, но начало истории человечества.
Конечно, на деле первый вал и второй не были строго разделены во времени: первый в одних географических областях еще продолжался, когда в других началось и зашло далеко встречное или обратное движение. Земля начала покрываться антропосферой: соприкасающимися друг с другом, но разделенными друг от друга первобытными образованиями. Земной шар перестал быть открытым для неограниченных перемещений. Его поверхность стала уже не только физической или биогеографической картой, но картой этногеографической, а много позже и политико-географической.
Единственное, что нас здесь касается в характеристике этих образований: они в общем всегда эндогамны. Этнос или другой тип объединения людей служит препятствием (иногда это – строгая норма, иногда – обычай, иногда – статистическая реальность) для брачно-половых связей с чужими. В таком трансформированном виде воспроизвелась внутри мира неоантропов биологическая инерция предшествовавшей дивергенции неоантропов с палеоантропами. Ведь несомненно, что к главнейшим механизмам дивергенции принадлежало избегание скрещивания (как показала этология, инстинкты, препятствующие скрещиванию, многообразны, даже у самцов и самок одного и того же вида они образуются на разной основе). Таким образом, эндогамия, разделившая мир неоантропов на взаимно обособленные ячейки, сделавшая его сетью этносов, была наследием дивергенции, как бы возведенным в степень, получившим совершенно новую функцию.